2025年11月7日,北京大学考古文博学院“宿白讲座”第七讲在北京大学考古楼A座101室举办,英国国家学术院院士、英国伦敦大学学院申南(Stephen Shennan)教授应邀作题为“文化与人口的计算建模:从理论到实践”(Computational Modelling of Culture and Demography: From Theory to Practice)的学术专题讲座。讲座由北京科技大学科技史与文化遗产研究院教授刘思然主持。

Stephen Shennan教授主讲
一、绪言:我们为何进行建模?
模型是对现实世界的简化表达,以实现对现实世界的预测。利用建模方法研究考古问题,有两个主要目的,第一是推断,基于证据与推理得出观点或结论,第二是识别反直觉的现象或涌现(emergent),当多个过程(或多个主体)发生互动时我们的直觉常常是错误的,计算建模可以帮助我们找到这些反直觉的或涌现的现象。在讲座中,申南教授通过两个案例对这两个应用场景分别进行说明。
涌现指复杂整体呈现出其组成部分单独不具备、仅在更大系统中相互作用时才出现的性质或行为。例如:鸟、鱼和蚂蚁的结群都由简单的行为规则产生,个体尽快回家的想法可能催生交通拥堵,简单化学过程可以催生复杂的生命系统。
对于这样的系统,我们能够理解其中个体的行为,但却很难预测系统的整体模式。为考察复杂系统行为的涌现,可以使用基于“主体”(agent)的模拟。它的主要思路是:首先建立大量可自主行动的主体,它可以是个人、家庭或者更大的群体;之后模拟主体的行为互动,主体与他者及环境的互动会遵循一定规则;对互动过程不断迭代,并在迭代达到一定时限后得到模拟结果;改变模型中的参数,观察参数变化对模拟结果的影响,进而产生规律性的认识。
二、狩猎采集群体的流动性建模
申南教授所举第一个案例是对狩猎采集群体的流动性建模,体现了反直觉现象的识别,研究成果已由申南教授与合作者发表(Gallagher et al., 2019)。狩猎采集人群产生流动性的主要原因是为了寻找更多食物,因为他们对某一地点资源的持续掠取会导致采集区资源的枯竭。此外,宗教活动、维系亲属关系、贸易、艺术的追求、个人义务,维系社会网络、择偶等也会增加人群流动性。然而,提升流动性会带来更多的能量消耗,因为流动过程需要群体进行额外的计划,同时也导致群体被掠食者袭击的风险升高,人群能够携带工具总量下降,丧失原采集区内的潜在机会,关于狩猎采集区的地方性知识降低等问题。那么,导致人群选择高或低流动性策略的影响因素具体有哪些?通过简单的对比可知,食物资源的类型、采集狩猎区的环境、人口规模都与流动性的高低具有相关性。
本研究采用基于主体建模的方法对多变量的协同作用开展研究,考察了主体移动频率与环境条件、适存度(可理解为群体的食物供给充足程度)及主体与环境的互动方式间的关系。每个主体都有各自的“流动性策略”,即其在单个时间步内从一地迁往另一地的概率,主体可以从他人处学习或通过试验改变自己的流动性策略。影响主体流动性策略的环境因素被定义为自然资源的觅食质量,它与食物资源的再生速率及狩猎采集人群对于食物资源的消耗速率有关。另一个重要因素是裂变概率,即一个主体分裂为更多主体的概率。当适存度高时,主体就会分裂产生新的主体,而当适存度过低时主体无法分裂并可能最终消亡。适存度会同时影响流动性策略的均值和分布,当适存度低时,流动性策略的均值和方差均较大,导致主体流动性的不确定性较高。反之,在适存度较高的情况下,流动性较低且确定性较高。
为了进一步定量化地描述这些问题,本研究在建模中设计了一系列参数以阐述以上机制的概率分布和相互关系,具体参数设计如图1所示。

图1 模型参数
在初始化主体的分布后,模型的迭代包含以下步骤:首先更新环境的觅食质量数据,这会导致部分主体消亡,部分主体发生分裂,产生新的主体;之后主体调整自身的流动性策略,进而发生流动;根据新地点的觅食质量数据,又有部分主体消亡;最后再次更新觅食质量数据,重复以上步骤。研究中建立了一个由10×10个六边形斑块组成的网格,作为模型运作的区域,用六边形斑块的颜色的深浅代表觅食质量的高低,建立模型可视化的表达,斑块中的数字表示其中主体的变化(图2)。
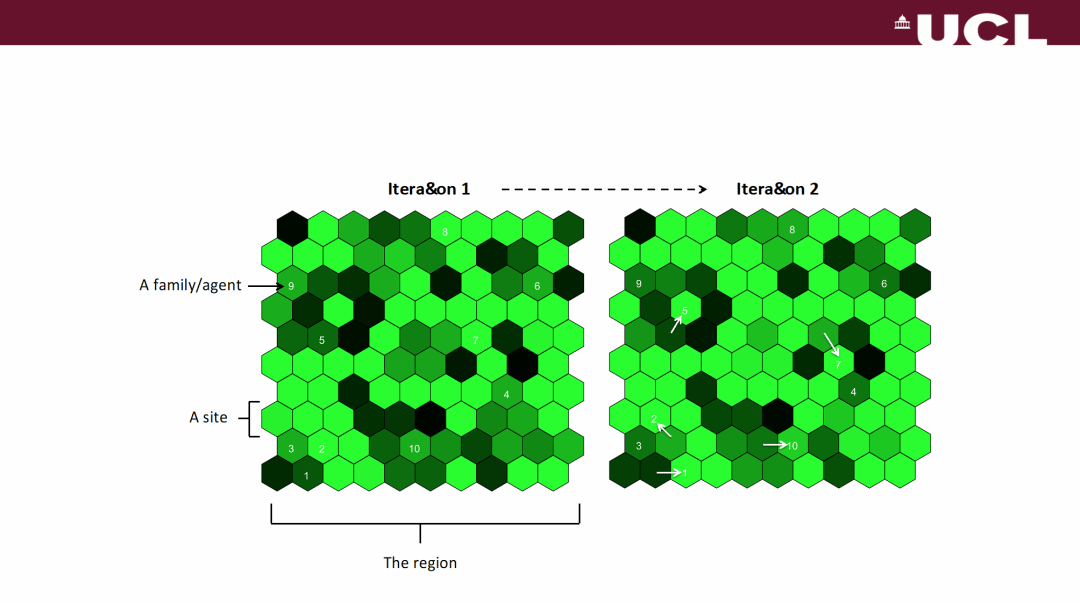
图2 模型可视化
使用两种环境条件下的模拟结果进行说明。首先是在低觅食质量再生率/高消耗条件下,资源易被耗尽且/或恢复缓慢。在这种环境下,主体的流动性策略会迅速提升并保持在很高的水平(蓝线),而主体数量则在低位徘徊(黑线),没有随时间增长,觅食质量与适存度则在中间区域波动(图3)。
第二次模拟是在环境具有高觅食质量再生率/低消耗条的条件下,此时资源不易被耗尽且/或恢复迅速。模拟结果显示,主体数量逐渐上升且维持在了较高水平,而主体的流动性策略取值则不断下降,显示定居化的倾向。值得注意的是,虽然此条件下觅食质量保持了很高的水平(绿线),但适存度却相对偏低(红线)(图4)。
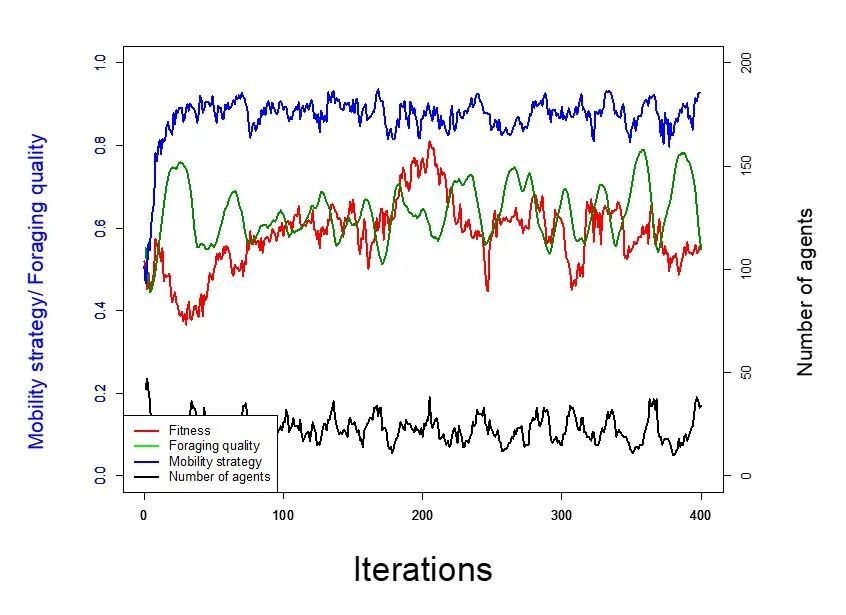
图3 低觅食质量再生率/高消耗
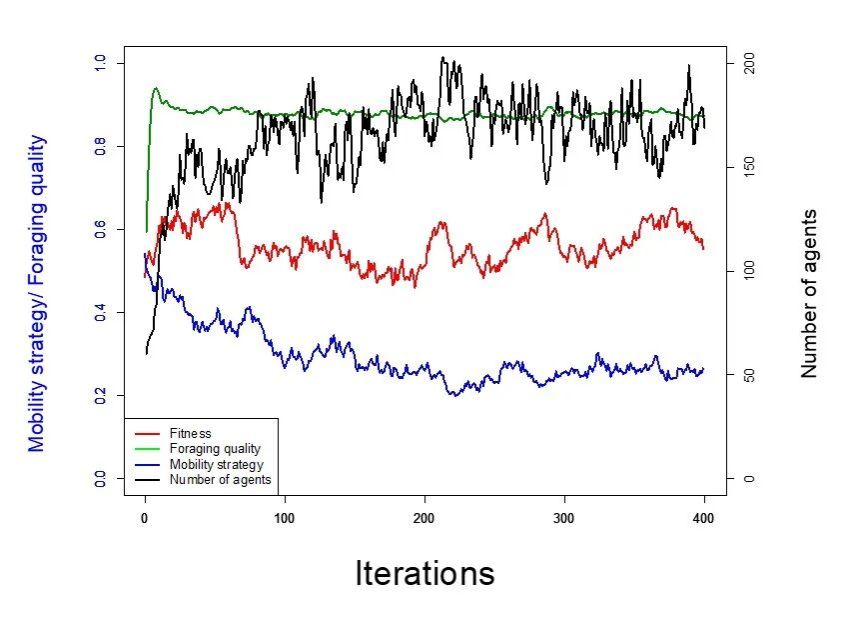
图4 高觅食质量再生率/低消耗
利用相同方式,研究者开展了10万次不同参数的模拟,并分析了参数与结果之间的相关性。结果显示,觅食质量消耗系数与流动性策略指示呈正相关,说明环境较差时人群更倾向于流动,反之则倾向于定居;主体数量与流动性策略指数呈负相关,说明更加定居的人群人口数量一般较高(图5)。这些观察与我们既往对狩猎采集人群流动性的认识相符。

图5 参数与结果的相关系数
然而,我们同时观察到最终环境适存度与最终主体数量、流动性策略指数存在强负相关关系,即人口越多、流动性越低的人群人均可获取的食物越少,生活质量越低。这一结果与我们以往认为生存在较好环境中的定居人口会有更高生活质量的认识的直觉相反。另一方面模拟结果显示,生活在觅食质量较差环境中的高流动性人群的食物质量较高。
人口由高流动性向定居化的转变与农业的出现息息相关。以往研究将这一转变带来的人群健康水平下降归因于食物质量/营养多样性下降,以及人口密度增加和城市化生活方式导致的病原体含量增加(Diamond, 2002)。然而,本次模拟研究的结果表明,即使不依靠这些外部因素,人群健康水平下降也是可解释的。产生这一现象的本质原因是,人口数量的自然增长与环境约束下的自然选择。
生活在较差环境中的高流动性人群,面临更加严苛的自然选择,他们适存度的不确定性更高,波动性更强,这导致当他们适存度均值较低时,一次较大的随机波动就会导致人群消亡,能够持续生存到模拟结束的人群往往具有较高的适存度均值。反之,生活在较好环境中的定居人群适存度波动较小,他们可以容忍较低的适存度均值,即较低的生活质量。而较高的觅食质量和较低适存度的协同作用(人口数量=觅食质量/适存度)使得定居人群的人口数量远高于流动人群。
三、最后的狩猎采集者与最初的农业人群之间的人口互动建模
早期农业人群的扩散是考古学界非常关心的问题。以往研究的视角更多关注农业人口的扩散速率、环境变化对农业的影响、记录狩猎采集人口和农业人口的时空分布、区分两群人的DNA等。这些研究的缺憾是没有关注这两个人群互动中的人口变化,不断扩张的农业人口如何影响原先居住于此的狩猎采集人口,反之又如何?这正是第二个案例所要解决的问题,此研究已由申南教授与合作者发表(Cortell-Nicolau. et al., 2025)。
在这个案例中,建模发挥了另一种作用——推断,当我们想知道原来设计的不同影响机制哪些发生了决定性作用时,可以通过数据与模型的拟合,获得各参数的后验分布,进而评价各类机制的作用程度。这个模型的核心问题是不断扩张的农业人口与既有的狩猎采集人口存在怎样的相互影响。首先使用遗址的碳十四年代汇总概率分布(SPD)估计两类人口随时间变化的情况,进而构建狩猎采集者与农业人口的互动模型以解释这些数据。
模型的框架使用洛特卡-沃尔特拉竞争模型,这是一对相关的微分方程,每个方程刻画一个群体的人口动态,并包含与另一群体相互作用所产生的影响。模型的两个关键要素是种内(群内)竞争与种间(群间)竞争,可用各自种群的内部增长率、群间的死亡率(竞争导致)和群间的转化率(狩猎采集向农业转化)来进行描述(图6)。另外模型的初始条件,即两类人群的初始比例,对于模拟结果的影响很大。这个取值的变化,可能导致导致不同的模拟结果,包括两类群体(长期或短期)稳定共存,或一方/双方消失。

图6 模型参数
为简化起见,本研究参照现有的考古文献材料设定了两类人群的初始比值,并重点对考古与民族志案例中最常见的情形(例如两群体先共存,随后狩猎采集人群逐渐消失)进行参数探索与实证检验。在模拟起始时设定狩猎采集人群的人口已处于环境承载力的最大值,而农业人群人口的环境承载力可随时间提高。这一设定符合两组人群的碳十四人口曲线经验观察结果。
首先使用与案例研究样本数量相当的模拟数据检验模型能否回收真实参数,以评估模型可靠性。拟合时采用近似贝叶斯计算(ABC)估计,使模型参数组合生成的碳十四数据汇总概率密度分布尽可能贴合考古记录,进而估计各参数的后验分布。结果显示,模型可以很好地回收产生模拟数据时所设定的参数。
本研究选取了丹麦,伊比利亚东部、日本九州三个区域的考古数据进行案例研究。每个区域建立两条碳十四的累计概率密度曲线,分别对应狩猎采集人群和农业人群的人口变化。将模型得出的最优拟合人口动态转化为碳十四年代汇总概率密度分布,显示三个区域的拟合效果均较好。三个区域的农业人群与狩猎采集人群人口关系显示出明显差异,丹麦的狩猎采集人群数量在初始状态下非常高,在与农业人口发生互动后,发生了迅速的衰减,而农业人口快速上升。日本初始的狩猎采集人群的数量并不多,农业人群人口却上升缓慢,与狩猎采集人群有很长的共存区间,之后随着狩猎采集人群的逐渐消亡,农业人群人口才开始快速上升(图7)。
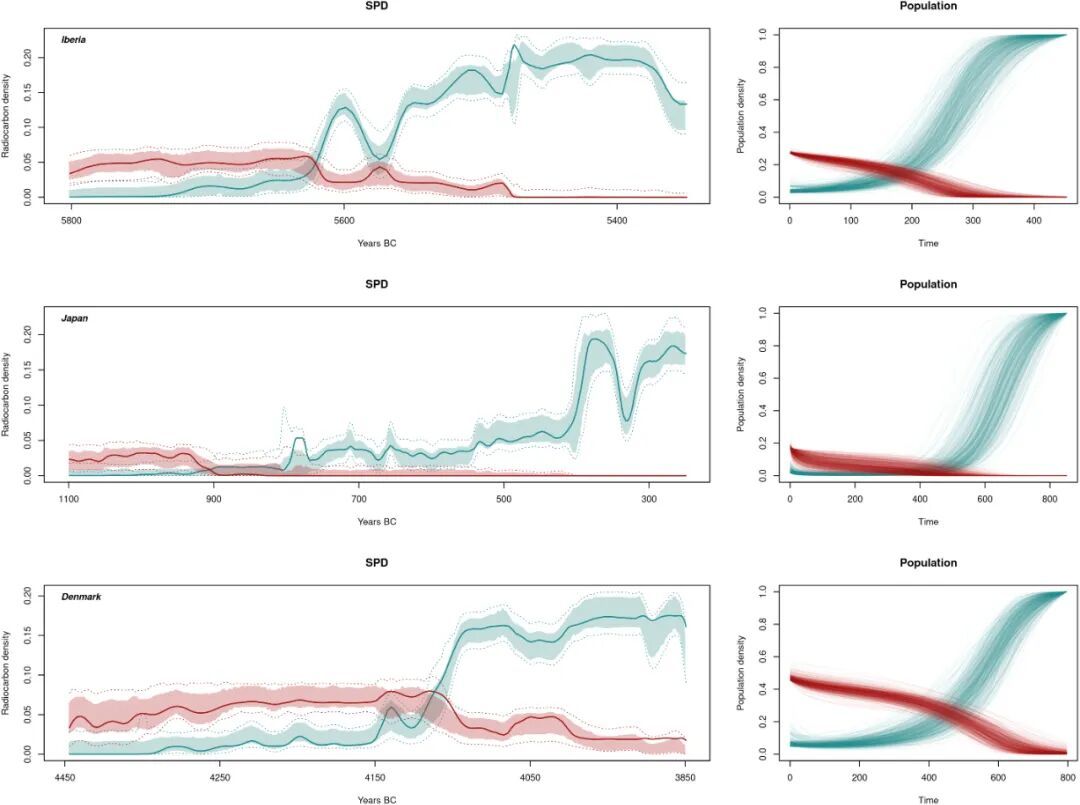
图7 最优拟合人口动态以及碳十四年代汇总概率密度分布
各参数的后验分布在几个区域间也有较明显的区别。比如丹麦和伊比利亚半岛东部的农业人口内增长率γf和群间转化率η明显高于日本,说明日本地区狩猎采集人群很少转化为农业人群,显示了该地区农业传播机制的特殊性(图8)。
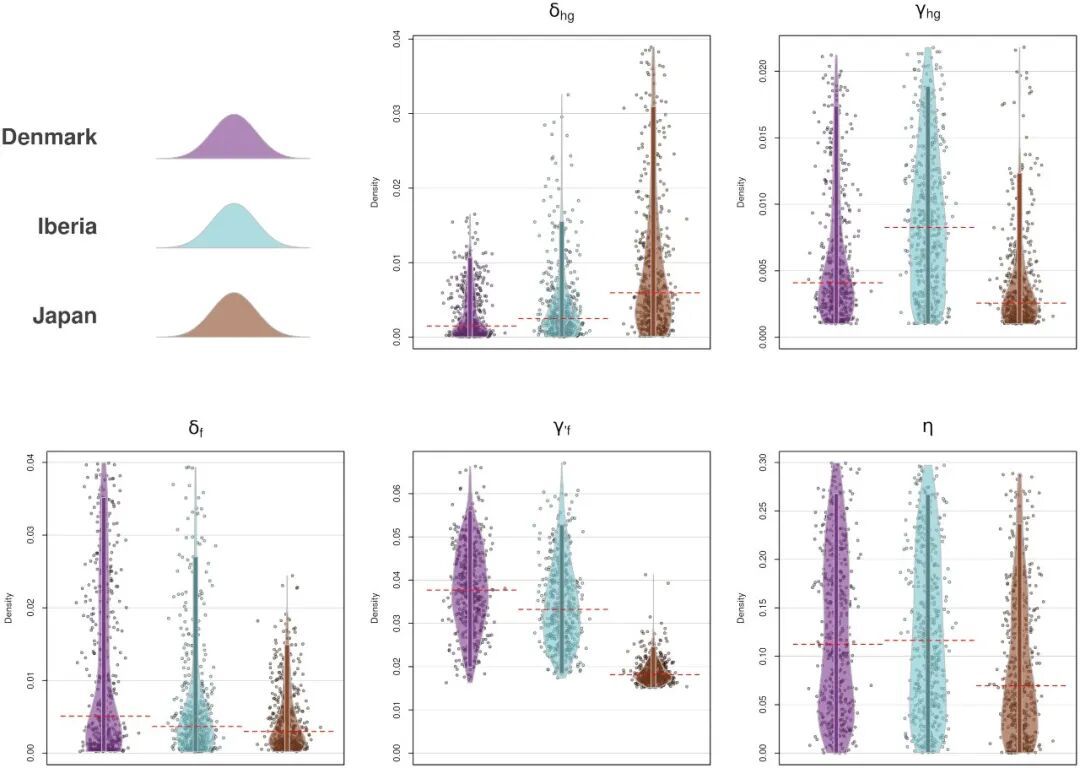
图8 参数后验分布
利用模型参数的后验概率分布还可计算各地区农业人群取代狩猎采集人群所需的时间长度。农业人群人口超过本地的狩猎采集人群人口所需时间以及狩猎采集人群完全消失所需时间的概率分布如图9所示,各地区间的差异也十分明显。
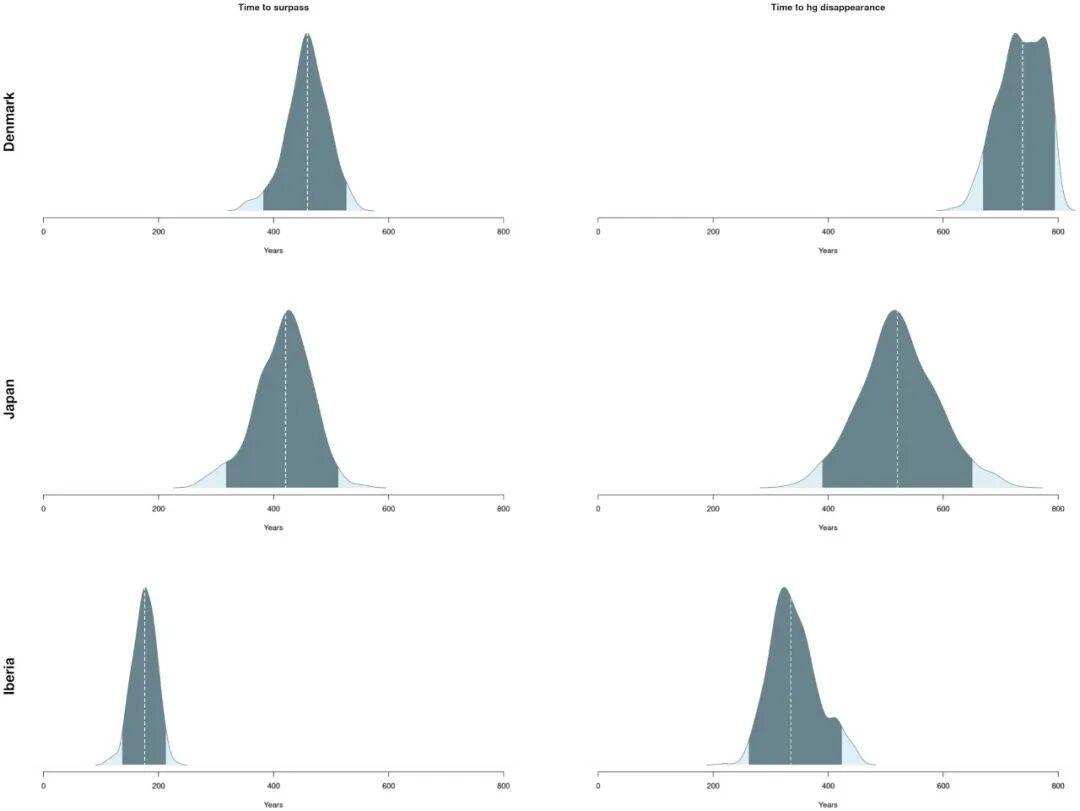
图9 取代过程的时间范围
这一模型对于研究广义的人口竞争具有重要意义,可用于探索本地群体与迁入群体之间多种类型的互动机制。将该理论模型与基于模拟的生成推断结合时,可对经验数据进行有效拟合。通过农业扩散的三则考古个例展示了此方法的潜力,并揭示了农业转型过程中人口增长与竞争对塑造人口变化轨迹所发挥的作用。
四、复杂社会问题的模型
以上两个模型都是对真实情况的高度简化,当遇到更加复杂的情况,如何对那些更加微妙的条件差异进行建模,则是这类型研究的关键挑战。由于人类社会的真实情况非常复杂,很多学者认为更复杂的模型才更加真实。然而,复杂也意味着引入更多参数,这给模型提供了更多偏离真实的可能性(自由度)。除非获得确切的相关数据,否则复杂的模型一般更容易出错,而非更加真实。如何通过模型阐明个体层面的、微观的、多样的互动对群体的宏观影响(例如对群体迁徙,扩张,遗传-文化关系、经济可持续性以及死亡年龄的影响),是建模工作的核心挑战。
申南教授简单展示这个方面的一项案例。研究者对文献现象如何通过通婚进行传播进行了模拟研究,探讨了婚后居住规则、性别偏见、移民者的文化接受度等因素对某项文化因素传播的影响(Carrignon et al., 2024)。其中一个关键的概念是身份信号与同质性现象。个体倾向与那些同自己更相似的人交往,这有可能促进群体内部的文化规范得到巩固,进而增加不同文化特征之间的关联,并在考古记录中形成界定清晰的物质文化群体。同质性的形成与文化传播的机制间有着紧密联系,当因婚姻发生迁徙的人口主要通过代际间学习获取文化身份信号时,文化的同质性会得到加强,而当这种学习主要在同代人间横向发生时,则会打破文化的同质性。
讲座最后,刘思然教授进行了总结:如何对数据进行科学有效的阐释,一直是考古学研究中的挑战。以往我们的大量工作停留在描述性层面,难以建立数据中观察到的现象与考古学阐释间的有效关联。申南教授在讲座中提出的建模方法可以有效的解决这一问题,我们可以将对于某个问题的构思以模型参数的方式体现出来,并通过模型拟合或生成去检验这些参数与观察数据间关联的显著性程度,更加科学化地思考如何对考古数据进行阐释。
参考文献
Carrignon S, Crema E R, Kandler A, et al. Postmarital residence rules and transmission pathways in cultural hitchhiking[J]. Proceedings of the National Academy of Sciences, 2024, 121(48): e2322888121.
Cortell-Nicolau A, Rivas J, Crema E R, et al. Demographic interactions between the last hunter-gatherers and the first farmers[J]. Proceedings of the National Academy of Sciences, 2025, 122(14): e2416221122.
Diamond J. Evolution, consequences and future of plant and animal domestication[J]. Nature, 2002, 418(6898): 700-707.
Gallagher E, Shennan S, Thomas M G. Food income and the evolution of forager mobility[J]. Scientific Reports, 2019, 9(1): 5438.


